Behind the paper: Four novel anti-CRISPR proteins found using synthetic biology
By EricvdHelm
Last week our article on new anti-CRISPR proteins was published in Cell Host & Microbe. Anti-CRISPRs are small proteins that inhibit the activity of CRISPR-Cas. These days a lot of research is focused on finding more active CRISPR-Cas systems. So why does anyone want to reduce CRISPR-Cas activity?
A big challenge with CRISPR-Cas is that it cleaves DNA where it is not supposed to, known as the “off-target effect”. This means CRISPR-Cas can introduce mutations that are unwanted. Unwanted mutations are just one of the many concerns about the recently edited “CRISPR babies”, as they could pose potential and unknown dangers to human health.
A solution to prevent off-target effects is to temporarily turn CRISPR-Cas activity off. This can be done using proteins that inhibit CRISPR-Cas activity. Some proteins mimic the DNA segment CRISPR-Cas is supposed to cut, while others prevent Cas9 from changing conformation. Until now, only a dozen anti-CRISPR proteins were known and are specific for their CRISPR-Cas type (see the figure below). So, it would be very useful to find more proteins that can inhibit CRISPR-Cas.

On the bottom the latest classification of CRISPR-Cas systems modified from Makarova and on the top the number of anti-CRISPR families found today as tracked by the anti-CRISPR database led by Joe Bondy-Denomy. In green, the anti-CRISPRs we found against Cas9. CRISPR-Cas systems are first divided into two classes based on which enzymes they use to cleave DNA or RNA sequences. In Class 1, this is actually a complex of several enzymes. In class 2, a single, but much larger enzyme is responsible for the final action. The layout of the tree is roughly based on the most conserved CRISPR-Cas gene, Cas1, and colored in deep purple.
The Idea
Back in 2015 I was working on detecting anti-HIV protease small molecules in metagenomic libraries using genetic circuits. Around that time genetic circuits were described that had a CRISPR-Cas output. Back then, CRISPR-Cas was not widely used in genetic circuits. We combined these concepts to find genes in metagenomic libraries that can inhibit a CRISPR-Cas system. Theoretically, it was possible because anti-CRISPRs against CRISPR-Cas type I-F and I-E were found in a ubiquitous bacteria, Pseudomonas aeruginosa. However, no anti-CRISPRs were known for use against the widely-used type II, better known as CRISPR-Cas9. This came in December 2016.
Until now, anti-CRISPRS were found computationally or by cloning out phage genes individually. We sought to perform the anti-CRISPR search in a high-throughput manner and without the prior knowledge that is needed for in silico prediction. The bacterial selection system we used contains two components, as outlined in the figure below.
The primary component is a genetic circuit with a Cas9 protein and guide RNA. This circuit cuts an antibiotic resistance gene on a plasmid, rendering the bacterial cells susceptible to antibiotics when no anti-CRISPR protein is present. The second component in the selection system is an anti-CRISPR source, in this case a metagenomic library. As input material we used metagenomic libraries that were constructed from fecal samples from humans, cows and pigs. Additionally we also used a metagenomic library from a soil sample. I previously wrote how similar systems are used to find new antibiotic resistance genes or vitamins transporters.
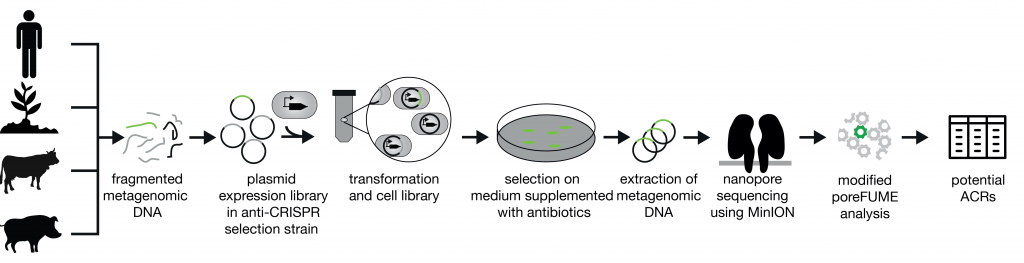
The discovery workflow starts with fecal samples from humans, cows and pigs, and a soil sample. It ends with a list of potential anti-CRISPR genes. The proteins encoded by the potential anti-CRISPR genes were then validated using various experimental methods.
The Experiments
After a lot of tweaking, colonies appeared that were able to dodge the selection system (and potentially Cas9 activity). Instead of sequencing individual colonies with Sanger sequencing, we used Nanopore sequencing. For Nanopore sequencing we used our previously published poreFUME protocol that allowed us to multiplex the sequencing of colonies and various different DNA sources.
The resulting sequences contained hits that did not only inhibit Cas9 activity, but also contained antibiotic resistance genes or genes that turned off expression of Cas9. After removing these false-postive genes, and to validate whether the sequences are actually inhibiting Cas9, the DNA was re-cloned into an E. coli expression vector. The individual proteins were tested in two ways: by assaying if Cas9 can still cleave DNA, and by testing whether the potential anti-CRISPR protein actually worked.
We visualized this process by putting the resulting reaction on a gel and checked whether the DNA was still intact, indicating that an anti-CRISPR protein was potentially inhibiting Cas9. If the DNA was cut into two pieces it was assumed the protein did not prevent Cas9 from cutting the DNA.
In the second experiment we tested with biolayer interferometry whether the potential anti-CRISPR protein can bind to Cas9. Together these experiments led us to believe that we found four new anti-CRISPR proteins, which we named AcrA7, AcrA8, AcrA9 and AcrA10. (Acr = anti-CRISPR. A because Cas9 is a type II-A system and 7-10 because of the chronological order they were found in, as proposed).
We unleashed a whole suite of computational analyses to find out how widespread these new anti-CRISPR proteins are. For example, we expected that the anti-CRISPR genes would co-occur with the CRISPR-Cas systems they are inhibiting, however there appeared no specific correlation between the two. A possible explanation could be that anti-CRISPR genes are often on mobile elements (such as plasmids and phages) and move through the population more rapidly than CRISPR-Cas systems. However, we did find homologs of the anti-CRISPRS in seven different phyla, including Firmicutes, Proteobacteria, Bacteroidetes, Actinobacteria, Cyanobacteria, Spirochaetes, and Balneolaeota, with high sequence similarity. This hints that the anti-CRISPR genes were moved recently by horizontal gene transfer.
Overall this was a very exciting research project, though we never optimized this selection system to find all of the possible anti-CRISPRs, nor did we analyze all the resulting clones extensively. We were initially just interested whether this setup could actually work to find new anti-CRISPR genes. I’m confident the anti-CRISPRs found thus far will eventually find their way into mainstream usage of CRISPR-based therapies and applications. Next, it will be exciting to look into more metagenomic libraries against all the other CRISPR types for which we don’t have a single anti-CRISPR yet. I’m sure they are out there!
Uribe, R. V, van der Helm, E., Misiakou, M., Lee, S., Kol, S., & Sommer, M. O. A. (2019). Discovery and Characterization of Cas9 Inhibitors Disseminated across Seven Bacterial Phyla Short Article Discovery and Characterization of Cas9 Inhibitors Disseminated across Seven Bacterial Phyla. Cell Host and Microbe, 25, 1–9. https://doi.org/10.1016/j.chom.2019.01.003
- ACR
- anti-CRISPR
- cell hostµbe
- CRISPR
- CRISPR-Cas
- functional metagenomics
- genetic circuit
- metagenomic library
- nanopore
- synbio
- synthetc biology